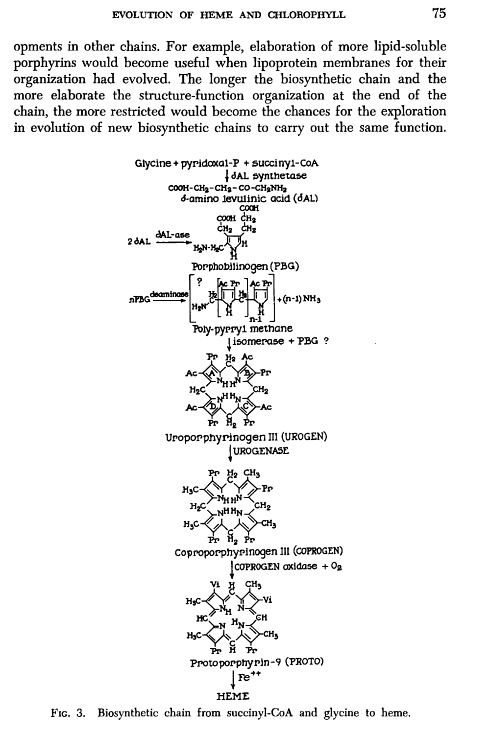
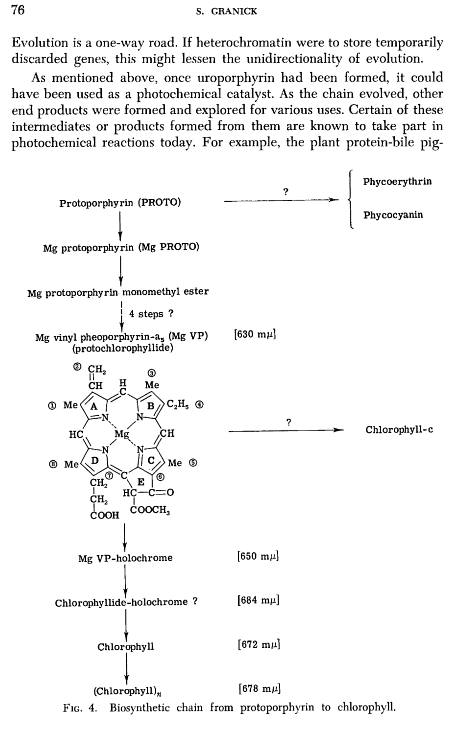
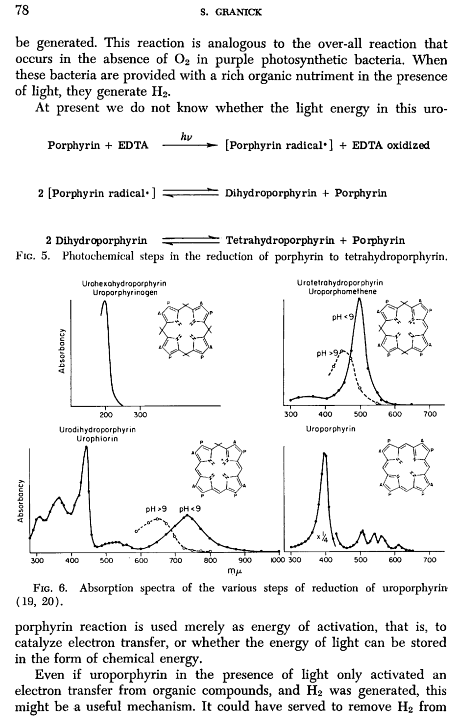
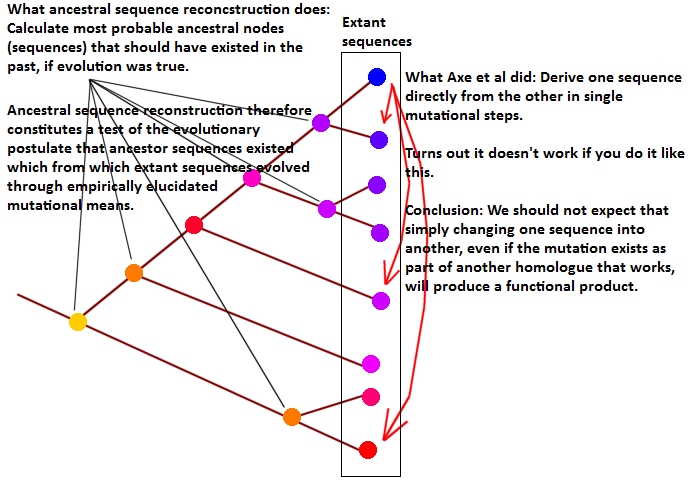
Dragan Glas
Well-Known Member
Greetings,
The improbable-odds-makes-it-impossible argument...
Given that you don't know what the odds are, simply assuming/claiming that "it's impossible" isn't a rational argument against evolution.
Kindest regards,
James
The improbable-odds-makes-it-impossible argument...
Given that you don't know what the odds are, simply assuming/claiming that "it's impossible" isn't a rational argument against evolution.
Kindest regards,
James